

| << Chapter < Page | Chapter >> Page > |
The Na +/ K + ATPase pumps in the basal membrane create an electrochemical gradient, allowing reabsorption of Cl – by Na + /Cl – symporters in the apical membrane. At the same time that Na + is actively pumped from the basal side of the cell into the interstitial fluid, Cl – follows the Na + from the lumen into the interstitial fluid by a paracellular route between cells through leaky tight junctions . These are found between cells of the ascending loop, where they allow certain solutes to move according to their concentration gradient. Most of the K + that enters the cell via symporters returns to the lumen (down its concentration gradient) through leaky channels in the apical membrane. Note the environment now created in the interstitial space: With the “back door exiting” K + , there is one Na + and two Cl – ions left in the interstitium surrounding the ascending loop. Therefore, in comparison to the lumen of the loop, the interstitial space is now a negatively charged environment. This negative charge attracts cations (Na + , K + , Ca ++ , and Mg ++ ) from the lumen via a paracellular route to the interstitial space and vasa recta.
The structure of the loop of Henle and associated vasa recta create a countercurrent multiplier system ( [link] ). The countercurrent term comes from the fact that the descending and ascending loops are next to each other and their fluid flows in opposite directions (countercurrent). The multiplier term is due to the action of solute pumps that increase (multiply) the concentrations of urea and Na + deep in the medulla.
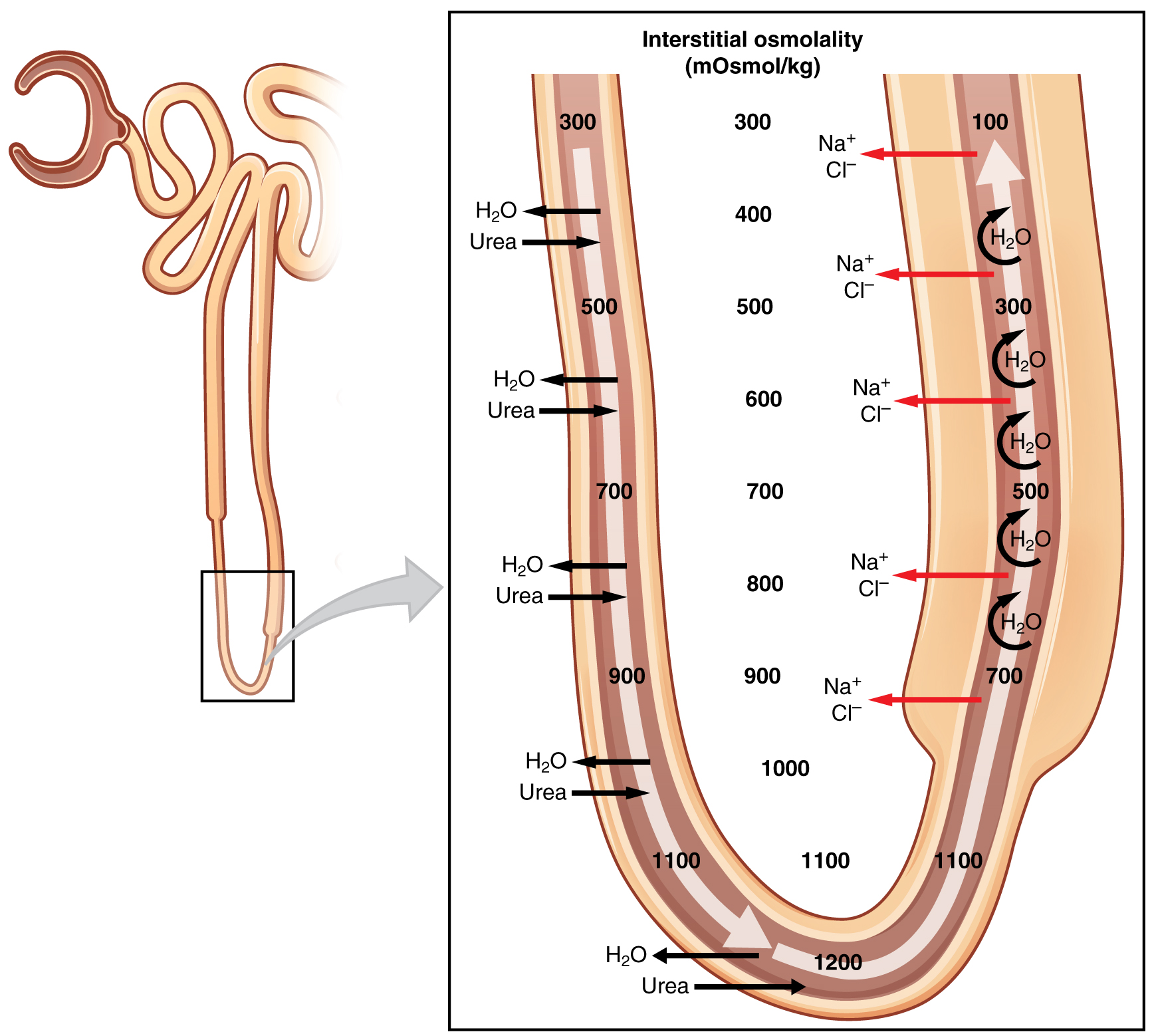
As discussed above, the ascending loop has many Na + pumps that actively pump Na + out of the forming urine into the interstitial spaces. In addition, collecting ducts have urea pumps that actively pump urea into the interstitial spaces. This results in the recovery of Na + to the circulation via the vasa recta and creates a high osmolar environment in the depths of the medulla.
Ammonia (NH 3 ) is a toxic byproduct of protein metabolism. It is formed as amino acids are deaminated by liver hepatocytes. That means that the amine group, NH 2 , is removed from amino acids as they are broken down. Most of the resulting ammonia is converted into urea by liver hepatocytes. Urea is not only less toxic but is utilized to aid in the recovery of water by the loop of Henle and collecting ducts. At the same time that water is freely diffusing out of the descending loop through aquaporin channels into the interstitial spaces of the medulla, urea freely diffuses into the lumen of the descending loop as it descends deeper into the medulla, much of it to be reabsorbed from the forming urine when it reaches the collecting duct. Thus, the movement of Na + and urea into the interstitial spaces by these mechanisms creates the hyperosmotic environment of the medulla. The net result of this countercurrent multiplier system is to recover both water and Na + in the circulation.
The amino acid glutamine can be deaminated by the kidney. As NH 2 from the amino acid is converted into NH 3 and pumped into the lumen of the PCT, Na + and HCO 3 – are excreted into the interstitial fluid of the renal pyramid via a symport mechanism. When this process occurs in the cells of the PCT, the added benefit is a net loss of a hydrogen ion (complexed to ammonia to form the weak acid NH 4 + ) in the urine and a gain of a bicarbonate ion (HCO 3 – ) in the blood. Ammonia and bicarbonate are exchanged in a one-to-one ratio. This exchange is yet another means by which the body can buffer and excrete acid. The presence of aquaporin channels in the descending loop allows prodigious quantities of water to leave the loop and enter the hyperosmolar interstitium of the pyramid, where it is returned to the circulation by the vasa recta. As the loop turns to become the ascending loop, there is an absence of aquaporin channels, so water cannot leave the loop. However, in the basal membrane of cells of the thick ascending loop, ATPase pumps actively remove Na + from the cell. A Na + /K + /2Cl – symporter in the apical membrane passively allows these ions to enter the cell cytoplasm from the lumen of the loop down a concentration gradient created by the pump. This mechanism works to dilute the fluid of the ascending loop ultimately to approximately 50–100 mOsmol/L.
Notification Switch
Would you like to follow the 'Anatomy & Physiology' conversation and receive update notifications?

|
|
|
|
|

|
|

|
|

|
|

|
|

|
|
|
|
|
|
|
|

|
|

|
|

|